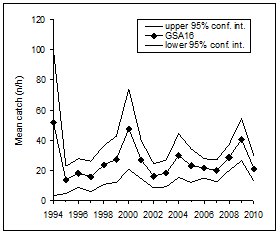
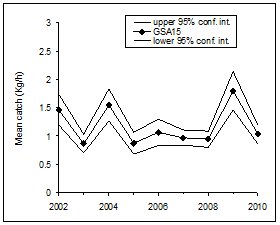
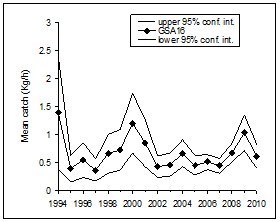
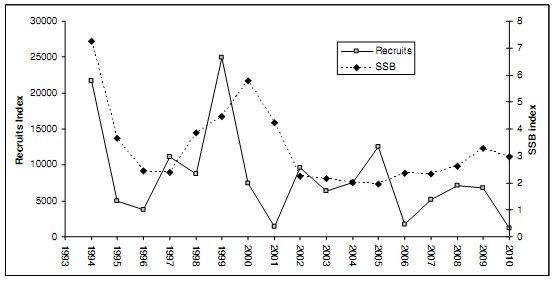
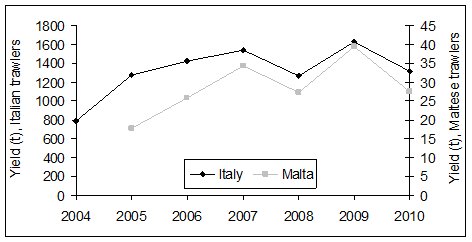
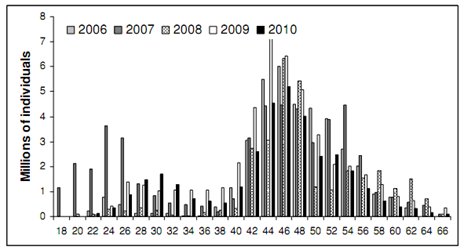
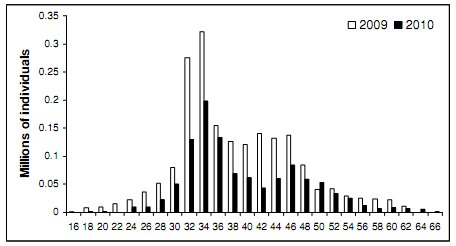
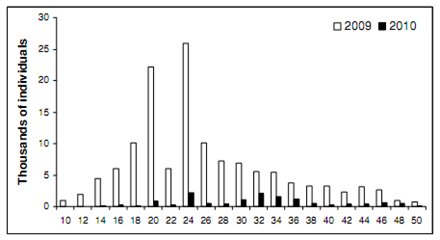
|
A.A.V.V. (2008). Status of deep-sea red
shrimps in the Central and Eastern Mediterranean Sea. Final Report. Project Ref.
FISH/2004/03-32.
Abelló P., Valladares F.J., Castellón A.
(1988). Analysis of the structure of decapod crustacean assemblages off Catalan
coast (North-West Mediterranean). Mar. Biol., 98: 39-49.
Abellò P., Abella A., Adamidou A.,
Jukic-Peladic S., Maiorano P., Spedicato M.T. (2002). Geographical patterns in
abundance and population structure of Nephrops norvegicus and Parapenaeus
longirostris (Crustacea: Decapoda) along the European Mediterranean coasts. Sci.
Mar., 66 (2): 125-141.
Alverson D.L., Freeberg S.S., Muraski S.S.,
Pope J.G. (1994). A global assessment of fisheries by-catch and discards. FAO
Fish. Tech. Pap., 339: 233 pp.
Andreoli, M.G., Campanella, N., Cannizzaro,
L., Garofalo, G., Giusto, G.B., Jereb, P., Levi, D., Norrito, G., Ragonese, S.,
Rizzo, P., Sinacori, G. (1995) Sampling statistics of southern Sicily trawl
fisheries (MINIPESTAT): data report. NTR–ITPP Special Publication 4 (vol. II).
Anon. (2000). Analysis of trawls’ discard
operation in the central and eastern Mediterranean Sea. Commission of the
European Communities. Directorate-General for Fisheries XIV. Contract No
97/0044. Final Report.
Ansell A.D., Robb L. (1977). The spiny
lobster Palinurus elephas in Scottish water. Mar. Biol., 43: 63-70.
Ardizzone G.D., Gravina M.F., Belluscio A.,
Schintu P. (1990). Depth–size distribution pattern of Parapenaeus longirostris
(Lucas, 1846) (Decapoda) in the central Mediterranean Sea. J. Crustac. Biol.,
10(1): 139-147.
Audouin J. (1965). Répartition bathymétrique
des crevettes sur les côtes algériennes entre les îles Zaffarines et les îles
Habibats. Commission Internationale pour l’Exploration Scientifique de la Mer
Méditerranée, 18: 171-174
Azouz A. (1971). Etude des biocénoses,
benthiques et de la faune ichtyologique des fonds chalutables de la Tunisie;
régions nord et sud-est. Thèse Sci. nat, Univ. Caen: 243 pp.
Azouz A. (1972). Les crustacés comestibles (crevettes
et langoustines) des mers tunisiennes. Bull. Int. natn. scient. tech. Océanogr.
Pêche. Salammbô, 2 (3) : 275 - 301.
Azouz A. (1981). Réflexions en vue de l’amélioration
de la production de la crevette royale dans le golfe de Gabès. Bull. Off. Nat.
Pêche. Tunisie, 5 (2) : 101 – 113.
Babbucci M., Buccoli S., Cau A., Cannas R.,
Goñi R., Díaz D., Marcato S., Zane L., Patarnello T. (2010). Population
structure, demographic history, and selective processes: contrasting evidences
from mitochondrial and nuclear markers in the European spiny lobster Palinurus
elephas (Fabricius, 1787). Mol. Phyl. Evol., 56: 1040-1050.
Bailey N. (1984). Some Aspects of
Reproduction in Nephrops. Shellfish Committee Document CM 1984/K:33. ICES: 16
pp.
Bauer R.T. (1991). Sperm transfer and storage
structures in penaeoid shrimp: a functional and phylogenetic perspective. In:
R.T. Bauer and J.W. Martin, Crustacean sexual biology, Columbia University
Press: 183-207.
Becer Özvarol Z.A., Tasly A., Ozvarol Y.
(2006). Some biological properties of the shrimp Metapenaeus monoceros (Fabricius,
1798) in the Gulf of Anatalya in the Mediterranean (Turkey). J.World. Aquac.
Soc.: 15 pp.
Belcari P., Viva C., Mori M., De Ranieri S.
(2003). Fishery and biology of Aristaeomorpha foliacea (Risso, 1827) (Crustacea:
Decapoda) in the Northern Tyrrhenian Sea (Western Mediterranean). J. Northw. Atl.
Fish. Sci., 31: 195-204.
Bell M.C., Redant F., Tuck I. (2006). "Nephrops
Species". In Bruce F. Phillips. Lobsters: Biology, Management, Aquaculture and
Fisheries. Wiley-Blackwell. 412–461.
Bello G., Pipitone C. (2002). Predation on
cephalopods by the giant red shrimp Aristaeomorpha foliacea. J. Mar. Biol. Ass.
U.K., 82(2): 213-218.
Ben Abdallah O. (2005). Contribution à l’étude
biologique et dynamique de la crevette mouchetée Metapenaeus monoceros (Fabricius,
1798) dans le golfe de Gabès (Tunisie). Mastère. INAT. 68 p.
Ben Abdallah-Ben Hadj Hamida O. (2012). Etude
biologique et dynamique de la crevette mouchetée Metapenaeus monoceros (Fabricius,
1798) exploitée dans le golfe de Gabès (Tunisie). Thèse de doctorat. INAT. 281
p.
Ben Hadj Hamida-Ben Abdallah O., Ben Hadj
Hamida N., Jarboui O., Fiorentino F., Missaoui H. (2009). Reproductive biology
of the speckled shrimp Metapenaeus monoceros (Fabricius, 1798) (Decapoda:
Penaeidae) in the gulf of Gabes (Southern Tunisia, Eastern Mediterranean).
Cahiers de Biologie Marine, vol. 50, no3, pp. 231-240.
Ben Hadj Hamida-Ben Abdallah O., Ben Hadj
Hamida N., Jarboui O., Missaoui H. (2010). Age and growth of the speckled shrimp
Metapenaeus monoceros (Fabricius, 1798) in the Gulf of Gabes (Southern Tunisia,
Central Mediterranean). Cahiers de Biologie Marine, vol. 51, no3, pp. 265-274.
Ben Khemis L. (1984). Pêche à la crevette
dans le golfe de Gabès. Rapp. Doc. Inst. natn. scient. tech. Océanogr. Pêche.
Salammbô, 2: 3 - 30.
Ben Meriem S. (1992). Eléments en vue d’un
aménagement des pêcheries du golfe de Gabès, Tunisie. Bull. Inst. natn. scient.
tech. Océanogr. Pêches. Salammbô, 19 : 66 - 84.
Ben Meriem S. (1993). Taille de première
maturité sexuelle et période de ponte de Penaeus kerathurus du golfe de Gabès (Decapoda,
Penaeoidea). Crustaceana, 65(1): 82 - 96.
Ben Meriem S. (1994). Aristaeomorpha foliacea
and Aristeus antennatus in Tunisian waters. Proc. Int. Workshop on life cycles
and fisheries of red shrimps, N.T.R.-I.T.P.P. Sec. Publ., 3: 50.
Ben Meriem S. (1995). Caractères biométriques
de Penaeus kerathurus du golfe de Gabès (Decapoda, Penaeoidea). Crustaceana,
68(5): 583 - 596.
Ben Meriem S. (1998). Mortalités (F et M) et
analyse des rendements par recrue de Penaeus kerathurus (Forskål) du golfe de
Gabès, Tunisie. Cah. Option Mediterr., 35, Dynamique des populations marines,
ISSN : 1022 – 1379 : 25 - 34.
Ben Meriem S., Fiorentino F., Arneri E., Ceriola L., Gancitano V., Jarboui O.,
Micallef R., Mifsud R. (2013). Stock assessment of pink shrimp (Parapenaeus longirostris) in GSAs 12-16. GFCM stock assessment form submitted at the GFCM
demersal stock assessment meeting in Bar, Montenegro, 03-04 February 2014.
Ben Mariem S. (2004). First approach to the
growth of Penaeus kerathurus (Decapoda, Penaeidae) in the Gulf of Gabès,
Tunisia. Crustaceana, 77: 277-297
Ben Meriem S., Gharbi H. (1988). Analyse
préliminaire des résultats de chalutage du “HANNOUN” dans le golfe de Gabès (du
2 au 22 août 1988). Rap. Doc. Inst. natn. scient. tech. Océanogr. Pêche Salammbô,
3: 1 – 34.
Ben Meriem S., Fehri-Bedoui R., Gharbi H.
(2001). Size at maturity and ovigerous period of the pink shrimp Parapenaeus
longirostris (Lucas, 1846) in Tunisia. Crustaceana, 74(1): 39-48.
Ben Mustapha A. (1967). Observations
biologiques sur Penaeus kerathurus
Forskål et étude biométrique. Annales. Inst.
natn. scient. tech. Océanogr. Pêche. Salammbô, (XIII).
Bevacqua D., Melià P., Follesa M.C., De Leo G.A., Gatto M., Cau A. (2010). Body
growth and mortality of the spiny lobster Palinurus elephas within and outside a
small marine protected area. Fish. Res., 106: 543-549.
Bianchini M.L., Di Stefano L., Ragonese S. (1998). Daylight vs. night variations
in the red shrimp catches of the Strait of Sicily. Rapp. Comm. Int. Mer Médit,
35(2): 374-375.
Bianchini M.L. (1999). The deep-water red shrimp
Aristaeomorpha foliacea of the
Sicilian Channel: biology and exploitation. University of Washington Ph.D.
dissertation: 482+ 17p.
Bianchini M.L., Ragonese S. (2002). Influence of the fishing ground on the
estimates of growth parameters in the deep-water red shrimp (Aristaeomorpha
foliacea) of the Strait of Sicily. Biol. Mar. Medit., 9(1): 732-735.
Bianchini M.L., Ragonese S., Levi D. (2003). Management hypotheses to improve
yield-per-recruit and economic returns in the red shrimp (Aristaeomorpha
foliacea) fishery of Southern Sicily (Mediterranean Sea). J. Northw. Atl. Fish.
Sci., 31: 233-243.
Branch, T.A., Kirkwood, G.P., Nicholson, S.A., Lawlor, B., Zara, S.J. (2000).
Yield version 1.0, MRAG Ltd, London, U.K.
Brian A. (1931). La biologia del fondo a "scampi" nel Mare Ligure. V.
Aristaeomorpha, Aristeus ed altri macruri natanti. Boll. Mus. Zool. Anat. Comp.
R. Univ. Genova, 2 ( 45) : 1-6.
Brutto, S. L., Maggio, T., Arculeo, M. (2013). Isolation By Distance (IBD)
signals in the deep-water rose shrimp Parapenaeus longirostris (Lucas, 1846) (Decapoda,
Panaeidae) in the Mediterranean Sea. Mar. Envir. Res., 90: 1-8.
Cacaud P. (2002) Etude comparative sur la réglementation en matière de pêche
maritime dans les pays de la Méditerranée occidentale participant au projet
Copemed. Projet FAO COPEMED. FAO, Rome : 34pp.
Camilleri M., Drago A., Fiorentino F., Garofalo G., Gristina M., Bahri T., Massa
F. (2008) Spatial pattern of fisheries demersal resources, environmental factors
and fishery activities in GSA 15. Pilot Study. MedSudMed GCP/RER/010/ITA/MSM-TD-13.
MedSudMed Technical Documents, 13:105pp.
Campillo A. (1982). Premières données sur la pèche et la biologie de la
langouste de Corse, Palinurus elephas Fabricius. Quad. Lab. Tecn. Pesca, 3(2-5):
115-39.
Campillo A., Amadei J. (1978). Premières données biologiques sur la langouste de
Corse, Palinurus elephas Fabricius. Rev. Trav. Inst. Pêches Marit., 42: 347-373.
Campillo A., P.Y. Dremiere, B. Liorzou, Bigot J.L. (1991). Observations sur deux
crustacés profonds du Golfe du Lion, Aristeus antennatus (R.) et Nephrops
norvegicus (L.). Rep Sixth Techn. Consultation Gen. Fish. Coun. Mediterranean,
FAO Fish. Rep., 447: 298-313
Can M.F., Aktas M. (2005). A preliminary study on population structure and
abundance of Aristaeomorpha foliacea (Risso, 1827) (Decapoda, Natantia) in the
deep water of the Northeastern Mediterranean. Crustaceana, 78 (8): 941-946.
Cannizzaro L., Vitale S., Arculeo M., De Stefano G., Lumare L., Milazzo A.,
Salvo G. (2011). Stock assessement and management by restocking of Melicertus
kerathurus (Forskål, 1775) in the shallow coastal.
Cannizzaro L., Alagna A., Andreoli M.G., Gianformaggio N. (1991) Relazione
taglia–peso per alcuni pesci del Canale di Sicilia. NTR–ITPP 39:33pp.
Carpenter K.E., Niem V.H. (eds) (1988). FAO species identification guide for
fishery purposes. The living marine resources of the Western Central Pacific.
Vol. 2. Cephalopods, crustaceans, holothurians and sharks. Rome, FAO.: 687-1396.
Carpentieri P., Colloca F., Ardizzone G. (2005) Day–night variations in the
demersal nekton assemblages on the Mediterranean shelf-break. Estuar. Coast.
Shelf Sci., 63:577-588.
Cartes J.E. (1995). Diets of, trophic resources exploited by, bathyal Penaeoidan
shrimps from the Western Mediterranean. Mar. Freshwater Res., 46: 889-896.
Cartes J.E., Company J.B., Maynou F. (1994). Deep-water decapod crustacean
communities in the Northwestern Mediterranean: influence of submarine canyons
and season. Mar. Biol., 120: 221-230.
Castriota L., Campagnuolo S., Andaloro F. (2001). Shrimp trawl fishery by-catch
in the Straits of Sicily (central Mediterranean Sea). Scientific Council
Research Documents of the Northwest Atlantic Fisheries Organization, Serial No.
N4501, No 01/113.
Cau A., Deiana A.M., Mura M. (1987). Ecological observations on Aristaeomorpha
foliacea (Risso, 1987) (Decapoda, Penaeidae) in the mit-Western Mediterranean
Sea. Invest. Pesq., 51 (1): 456.
Cau A., Carbonell A., Follesa M.C., Mannini A., Orsi Relini L., Politou C.Y.,
Ragonese S., Rinelli P. (2002). MEDITS-based information on the deep water red
shrimps Aristaeomorpha foliacea and Aristeus antennatus (Crustacea: Decapoda:
Aristeidae). Sci. Mar., 66 (S2): 103-124.
Ceccaldi H.J., Latrouite D. (2000). The French fisheries for the European spiny
lobster Palinurus elephas. In: B.F. Phillips & J. Kittaka (Eds) Spiny Lobster
Fisheries and Culture (2nd edn). Blackwell Science, Oxford, : 200–209.
Chaouachi B., Ben Hassine O.K. (1998) Données sur la pêche des crevettes
profondes Parapenaeus longirostris (Lucas, 1846) en Tunisie. Cah. Options Medit.,
35: 201–213.
CNR–IAMC (2006). Programma Nazionale Italiano per la raccolta di dati alieutici.
Modulo “Altri campionamenti biologici”- Risorse demersali - Unità Gestionale 16
(Stretto di Sicilia): rapporto finale, CNR–IAMC, Mazara del Vallo (TP), Italy:
46pp.
CNR–IAMC (2007). Programma Nazionale Italiano per la raccolta di dati alieutici.
Modulo H “Campionamento biologico delle catture” (CAMP–BIOL; Risorse demersali)
- GSA 16 (Stretto di Sicilia): rapporto finale, CNR–IAMC, Mazara del Vallo (TP),
Italy: 186 pp.
CNR-IAMC (2009). Programma Nazionale Italiano per la raccolta di dati alieutici
2008. Modulo I. Altri campionamenti biologici risorse demersali e piccoli
pelagici - GSA 16 (Stretto di Sicilia): relazione finale, IAMC-CNR, Mazara del
Vallo (TP), Italia: 79 pp.
Colloca F., M. T. Spedicato, E. Massutí, Garofalo G., G. Tserpes, P. Sartor, A.
Mannini, A. Ligas, G. Mastrantonio, B. Reale, C. Musumeci, I. Rossetti, M.
Sartini, M. Sbrana, F. Grati, G. Scarcella, M. Iglesias, M.P. Tugores, F.
Ordines, L. Gil de Sola, G. Lembo, I. Bitteto, M.T. Facchinii, A. Martiradonna,
W. Zupa, R. Carlucci, M.C. Follesa, P. Carbonara, A. Mastradonio, Fiorentino F.,
Gristina M., Knittweis L., Mifsud R., Pace M.L., C. Piccinetti, C. Manfredi, G.
Fabi, P. Polidori, L. Bolognini, R. De Marco, F. Domenichetti, R. Gramolini, V.
Valavanis, E. Lefkaditou, K. Kapiris, A. Anastasopoulou and N. Nikolioudakis,
2013 Mapping of nursery and spawning grounds of demersal fish. Mediterranean
Sensitive Habitats (MEDISEH) Final Report, DG MARE Specific Contract SI2.600741,
Heraklion (Greece).
Company J.B., Maiorano P., Tselepides A., Politou C.-Y., Plaiti W., Rotllant M.,
Sardà F. (2004). Deep-sea decapod crustaceans in the western and central
Mediterranean Sea: preliminary aspects of species distribution, biomass and
population structure. Sci. Mar., 68 (3): 73-86.
Conides A., Glamuzina B., Dulcic J., Kapiris K., Jug-Dujakovic J.,
Papaconstantinou C. (2008). Study of the reproduction of the karamote shrimp
Penaeus (Melicertus) kerathurus in Amvrakikos Gulf, western Greece. Acta Adriat.,
49(2): 97 – 106.
Conides A., Glamuzina B., Jug-Dujacovic J., Papaconstantinou C., Kapiris K.
(2006). Age, growth, and mortality of the karamote shrimp, Penaeus (Melicertus)
kerathurus (Forskål 1775) in the east Ionian Sea (Western Greece). Crustaceana,
79(1): 33-52.
Cristo M., Cartes J.E. (1998). A comparative study of the feeding ecology of
Nephrops norvegicus (L.), (Decapoda: Nephropidae) in the bathyal Mediterranean
and the adjacent Atlantic. Sci. Mar., 62 (1): 81-90.
Crnković D. (1965). Ispitivanje ekologije i mogućnosti racionalnog unaprijeđenja
eksploatacije raka Nephrops norvegicus (L) u kanalskom području sjeveroistočnog
Jadrana. Disertacija, PMF Sveučilišta u Zagrebu.
Cuccu D. (1999). Biologia ed Ecologia dell’ Aragosta Rossa (Palinurus elephas,
Fabricius, 1787). Proposte Gestionali per il Recupero dello Stock. Tesi di
dottorato in biochimica comparata degli ecosistemi, Ciclo XIV, Università di
Sassari, 111pp.
Cuccu D., Follesa M.C., Secci E., Cau A. (1999). Preliminary data on the
movement, growth, mortality and tag retention of the spiny lobster (Palinurus
elephas). European crustacean conference.
Cunningham J.T. (1891). On the development of Palinurus vulgaris, the Rock
Lobster or sea crayfish. J. Mar. Biol. Ass. U.K., 2: 141-150.
Cushing D.H. (1968). Fisheries biology. A study in population dynamics.
University of Wisconsin Press, Madison, 200 pp.
De Ranieri S., Mori M., Sbrana M. (1998). Preliminary study on the reproductive
biology of Parapenaeus longirostris (Lucas) off the northern Tyrrhenian Sea.
Biol. Mar. Medit., 5(1): 710-712.
Díaz D., Marí M., Abelló P., Demestre M. (2001). Settlement and Juvenile Habitat
of the European Spiny Lobster Palinurus elephas (Crustaceana: Decapoda:
Palinuridae) in the Western Mediterranean Sea. Sci. Mar., 65(4): 347–356.
Dimech M., Kaiser M.J., Ragonese S., Schembri P.J. (2012). Ecosystem effects of
fishing on the continental slope in the Central Mediterranean Sea. Mar. Ecol.
Prog. Ser., 449: 41-54.
D’Onghia G., Maiorano P., Matarrese A., Tursi A. (1998). Distribution, biology
and population dynamics of Aristaeomorpha foliacea (Risso, 1827) (Decapoda,
Natania, Aristaeidae) in the North-Western Ionian Sea (Mediterranean Sea). Crustaceana 71 (5): 518-544.
D’Onghia G., Mastrototaro F., Matarrese A., Politou C., Mytilineou C. (2003).
Biodiversity of the upper slope demersal community in the eastern Mediterranean:
preliminary comparison between two areas with and without trawl fishing. J. Northw. Atlant. Fish. Sci., 31: 263.
d’Udekem d’Acoz C. (1999). Inventaire et distribution des crustace´s de´capodes
de l’Atlantique nord-oriental, de la Me´diterrane´e et des eaux douces
continentales adjacentes au nord de 25°N. Patrimoines Naturels (MNHN/SPN) 40:
1–383.
D’Onghia G., Matarrese A., Maiorano P., Perri F. (1998) Valutazione di
Parapenaeus longirostris (Lucas, 1846) (Crustacea, Decapoda) nel Mar Ionio. Biol.
Mar. Medit. 5(2):273-283.
Dos Santos A. (1998). On the occurrence of larvae of
Parapenaeus longirostris
off the Portuguese Coast. J. Nat. Hist., 32: 1519-1523.
Eggleston D.B., Lipcius R.N. (1992). Shelter selection by spiny lobster under
variable predation risk, social conditions, and shelter size. Ecol., 73:
992-1011.
Falciai L., Minervini R. (1996). Guide des homards, crabes, langoustes,
crevettes et autres. Crustacés Décapodes d’Europe. Delachaux et Niestlé SA,
Lausanne-Paris. 1-287 pp.
FAO FishStat Plus 2012. Fisheries and aquaculture software. FishStat Plus -
Universal software for fishery statistical time series. In: FAO Fisheries and
Aquaculture Department [online]. Rome. Updated 28 November 2013. Available at
the URL: http://www.fao.org/fishery/statistics/software/fishstat/en.
Fiorentini L., Cosimi G., Sala A., Palumbo V. (1994). Caratteristiche e
prestazioni delle attrezzature a strascico impiegate per la valutazione delle
risorse demersali in Italia. Biol. Mar. Medit., 1 (2): 115-134.
Fiorentino F., Bono G., Garofalo G., Gristina G., Ragonese M., Gancitano S.,
Giusto G.B., Rizzo P., Sinacori G. (2003). A further contribution on stock’s
status and fisheries of main demersal resources in the Strait of Sicily. EN/TN/FF-GB-GG-MR-SR-SG-GBG-PR-GS/4/0303/DRAFT.
Fiorentino F., Garofalo G., Gristina G., Gancitano S., Norrito G. (2004). Some
relevant information on the spatial distribution of demersal resources, benthic
biocoenoses and fishing pressure in the Strait of Sicily. In: Report of the
MedSudMed Expert Consultation on Spatial Distribution of Demersal Resources in
the Straits of Sicily and the Influence of Environmental Factors and Fishery
Characteristics. GCP/RER/010/ITA/MSM-TD-02. MedSudMed Technical Documents, 2:
50-66.
Fiorentino F., Mazzola S., Garofalo G., Patti B., Gristina M., Bonanno A., Massi
D., Basitone G., Cuttitta A., Giusto G.B., Gancitano S., Sinacori G., Rizzo P.,
Levi D., Ragonese S. (2005). Lo stato delle risorse demersali e dei piccoli
pelagici e le prospettive di pesca “sostenibile” nello Stretto di Sicilia.
Convenzione con Assessorato Regione Siciliana Cooperazione, Commercio,
Artigianato e Pesca, Mazara del Vallo, Italy. ID/TN/FF-SM-GG-BP-MG-AB-DM-GB-AC-GBG-SG-GS-PR-DL-SR/8/0305/rel.
1:136pp.
Fischer W., Bauchot M.L., Schneider M. Eds.
(1987). Fiches FAO d’identification des espèces pour les besoins de la pêche. (Révision
1). Méditerranée et mer Noire. Zone de pêche 37. Volume I. Végétaux et
Invertébrés. Publication préparée par la FAO, résultat d’un accord entre la FAO
et la Commission des Communautés Européennes (Projet GCP/INT/422/EEC) financée
conjointement par ces deux organisations. Rome, FAO, Vo1.1: 267-268.
Follesa M.C., Cuccu D., Damele F., Sabatini A., Cau A. (2003). Valutazioni
sull’accrescimento di Palinurus elephas (Fabr., 1787) tramite marcatura e
ricattura nei mari sardi. Biol. Mar. Medit., 10 (2): 253-256.
Follesa M.C., Cuccu D., Cannas R., Cau A. (2007). On the growth of the European
spiny lobster, Palinurus elephas from Sardinian waters (Central Western
Mediterranean sea). N.Z. J. Mar. Freshw. Res., 41: 377-383.
Follesa M.C., Cuccu D., Damele F., Sabatini A., Cau A. (2007). Emigration and
retention of Palinurus elephas (Fabricius, 1787) in a region of the Central
Western Mediterranean MPA. Sci. Mar., 71 (2): 279-285.
Follesa M.C., Cuccu D., Cannas R., Sabatini A., Deiana A.M., Cau A. (2009).
Movement patterns of the spiny lobster Palinurus elephas (Fabricius, 1787) from
a central western Mediterranean protected area. Sci. Mar., 73(3): 499-506.
Froglia C., Gramitto M.E. (1988). An estimate of growth and mortality parameters
for Norway lobster (Nephrops norvegicus) in the Central Adriatic Sea. FAO Fish.
Rep., 394: 189-203.
Gaamour A., Rjeibi O., Bdioui M., Missaoui H. (2005). Les pêcheries de la
langouste rouge Palinurus elephas de la région Nord de la Tunisie : engin de
pêche, éléments biologiques et gestion. Rapport final du projet de recherche sur
la langouste rouge dans le cadre des activités de recherche du projet COPEMED.
Publié dans le site du Projet COPEMED.
Gaamour A., Rjeibi O., Ben Salem S., Jaziri S. (2009). Analysis of
Palinurus
elephas fisheries of the North coasts of Tunisia. Proceedings of TJASSST’10,
edited by: A. Ghorbel, A. Ben Salem, S. Bouzayene, pp. 112-115.
Galhardo A.M., Serafim P., Castro M. (2006). Aspects of the biology and fishery
of the European spiny lobster (Palinurus elephas) from the southwest coast of
Portugal. J. Crust. Biol., 26: 601–609.
Galil B., Froglia C., Nöel P. (2002). CIESM Atlas of Exotic Species in the
Mediterranean. Vol. 2. Crustaceans: decapods and stomatops. [F. Briand, Ed.].
192 pages. CIESM Publishers, Monaco.
Gamulin T. (1955). Contribution à la connaissance de l´écologie de la langouste
(Palinurus vulgaris Latreille) dans l'Adriatique. Act. Adriat., VII, 3-17.
Gamulin T. (1955). Contribution à la connaissance de l’écologie de la langouste
(Palinurus vulgaris Latreille). Arch. Zool. Exp. Gén., 67: 32-39.
Gancitano V., Cusumano, S., Giusto, G. B, Garofalo, G., Ingrande, G., Sabatella,
E., Ragonese, S., Fiorentino, F. (2008). Valutazione dello stato di sfruttamento
del gambero rosso Aristaeomorpha foliacea (Risso, 1827) (Crustacea; Decapode)
nello Stretto di Sicilia. Biol. Mar. Medit., 15(1): 326-327.
Gancitano V., Garofalo G., Gristina M., Ragonese S., Rizzo P., Gancitano S.,
Cusumano S., Ingrande G., Badalucco C., Fiorentino F. (2011). Potential Yield
and Current Exploitation of Deep Water Pink Shrimp (Parapenaeus longirostris),
Hake (Merluccius merluccius) and Giant Red Shrimp (Aristaeomorpha foliacea) in
the Strait of Sicily. In: Brugnoli E., Cavarretta G., Mazzola S., Trincardi F.,
Ravaioli M., Santoleri R. (eds) Marine research at CNR - Fishery and Sea
Rresources. - ISSN 2239-5172 Volume DTA/06-2011, Roma, November 2011, pp. 1-18.
Garofalo G., Fortibuoni T., Gristina M., Sinopoli M., Fiorentino F. (2011).
Persistence and co-occurrence of demersal nurseries in the Strait of Sicily (central
Mediterranean): Implications for fishery management. J. Sea Res., 66: 29-38.
Garofalo G., Giusto, G. B., Cusumano, S., Ingrande, G., Sinacori, G., Gristina,
M., Fiorentino, F. (2007). Sulla cattura per unità di sforzo della pesca a
gamberi rossi sui fondi batiali del mediterraneo orientale. Biol. Mar. Medit.,
14(2): 250-251.
Gashout S., Haddoud D., Zgozi S. (1992) Distribution–abundance of commercial
shrimp in Libyan waters, data from 1970 surveys. Arab Fish. J., Vol. 13.
Gayanilo Jr. F.C., Soriano M., Pauly D. (1988). A draft guide to the COMPLEAT
ELEFAN. ICLARM Software Project 2: 65 p. and diskettes
Gayanilo Jr. F.C., Sparre P., Pauly D. (1994). The FAO-ICLARM Stock Assessment
Tools (FiSAT) User's guide. FAO Computerized Information Series (Fisheries) No.
8 Rome, FAO, 124 p. and 3 diskettes.
Gayanilo Jr. F.C., Sparre P., Pauly D. (2005). Food And Agriculture Organization
Of The United Nations – Fisat II (version 1.2.2) Rome, 2005.
George J.P. (1977). Prospection des lieux de pêche de la crevette
Penaeus
kerathurus et introduction d'une nouvelle technique de pêche pour ce crustacé en
Tunisie (1973-1975). Bull. Inst. natn. scient. tech. Océanogr. Pêches Salammbô,
4 (2-4): 263 - 306.
Gharbi A., Ben Meriem S. (1996). Résultats de chalutages expérimentaux (mai-juin
1996) dans le golfe de Gabès. Bulletin de l’Institut National des Sciences et
Technologies de la Mer, 23: 80-96.
Ghidalia W., Bourgeois F. (1961). Influence de la temperature et de l’eclairement
sur la distribution des crevettes des moyennes et grandes profondeurs. Stud.
Rev. Gen. Fish. Count. Medit., FAO: 16: 53 pp.
Giacalone V.M., D’anna G., Pipitone C., Badalamenti F. (2006). Movements and
residence time of spiny lobsters Palinurus elephas released in a marine
protected area: an investigation by ultrasonic telemetry. J. Mar. Biol. Ass.
U.K., 86: 1101-1106.
Gislason H., Daan N., Rice J.C., Pope J.G. (2010). Size, growth, temperature and
the natural mortality of marine fish. Fish Fish., 11: 149–158.
Goñi R., Hilborn R., Diaz D., Mallol S., Adlerstein S. (2010). Net contribution
of spillover from a marine reserve to fishery catches. Mar. Ecol. Prog. S., 400:
233-243.
Goñi R., Latrouite D. (2005). Review of the biology, ecology and fisheries of
Palinurus spp. species of European waters: Palinurus elephas (Fabricius, 1787)
and Palinurus mauritanicus (Gruvel, 1911). Cah. Biol. Mar., 46(2): 127-142.
Goñi R., Quetglas A., Reñones O. (2001a). Diet of the spiny lobster Palinurus
elephas (Decapoda: Palinuridea) from the Columbretes Islands Marine Reserve (north-western
Mediterranean). J. Mar. Biol. Ass. U.K., 81: 347-348.
Goñi R., Quetglas A., Renones O. (2006). Spillover of spiny lobsters
Palinurus
elephas from a marine reserve to an adjoining fishery. Mar. Ecol. Progr. Ser.,
306: 207-219.
Goñi R., Quetglas A., Reñones O., Mas, J. (2003c). Threats to the sustainability
of Palinurus elephas fisheries. The Lobster Newsletter, 16, 2–5.
Goñi R., Quetglas A., Reñones R. (2003d). Biología, ecología, pesquerías y
efecto reserva de la langosta roja, Palinurus elephas Fabricius 1787 de
Columbretes y Baleares (Mediterráneo Occidental). Report 03-01, 1-107. 2003.
IEO-COB/LANGOSTA.
Goñi R., Quetlas A., Reñones O. (2003a). Size at maturity, fecundity and
reproductive potential of a protected population of the spiny lobster Palinurus
elephas (Fabricius, 1787) from the Western Mediterranean. Mar. Biol., 143:
583-592.
Goñi R., Quetlas A., Reñones O. (2003b). Differential catchability of male and
female European spiny lobster Palinurus elephas (Fabricius, 1787) in traps and
terammelnets. Fish. Res., 65: 295-307.
Goñi R., Reñones O., Quetglas A. (2001b). Dynamics of a protected western
mediterranean population of the european spiny lobster Palinurus elephas (Fabricius,
1787) assessed by trap survey. Mar. Freshw. Res., 52: 1577–1587.
Goñi, R. (2006). Proyecto langosta. Biología, ecología, pesquerías y efecto
reserva de la langosta roja Palinurus elephas (Fabricius 1787) en el
Mediterráneo Español. Revista electrónica del IEO 4, 21-26. 2006.
Groffiths R.C., Robles R., Coppola S.R., Camiñas J.A. (2007). Is there a future
for artisanal fisheries in the western Mediterranean? FAO, Rome. 106pp.
Gristina M., Fiorentino F., Garofalo G., Ragonese S., Gagliano M. (2002). Gli
effetti della protezione sull'aragosta comune (Palinurus elephas Fabricius,
1787) nella riserva marina delle "Isole Egadi". Report Rapporto preliminare, ID/W.PR./MG-FF-GG-SR-MG/2/07/02/DRAFT,
1-23.. Mazara del Vallo (TP), Italy, Instituto di Ricerche sulle Risorse Marine
e l'Ambiente - Consiglio Nazionale delle Ricerche.
Gristina M., Fiorentino F., Garofalo G., Gagliano M., Morizzo G., Cusumano S.
(2005). Effetti della protezione sull'aragosta comune (Palinurus elephas Fabricius, 1787) nella Riserva Marina delle Isole Egadi. Biol. Mar. Medit., 12:
404-409.
Gristina M., Fiorentino F., Garofalo G., Sinopoli M. (2008). L’aragosta comune
Palinurus elephas (Fabricius, 1878), nell’Area Marina Protetta di Capo Gallo –
Isola Delle Femmine. Processi di reclutamento ed impatto della pesca. Pesca e
gestione delle aree marine protette, workshop Porto Cesareo, 30 - 31 ottobre
2008.
Gristina M., Sinopoli M., Fiorentino F., Garofalo G.,·Badalamenti F. (2011).
Shelter selection of the spiny lobster Palinurus elephas under different levels
of Octopus vulgaris predation threat. Mar. Biol., 158: 1331–1337.
Groeneveld J.C., Goni R., Latrouite D. (2006). "Palinurus species". In Bruce F.
Phillips. Lobsters: Biology, Management, Aquaculture and Fisheries. John Wiley &
Sons. pp. 385–411. ISBN978-1-4051-2657-1.
Guijarro B., Valls M., Massutí E. (2010). SAC GFCM - Sub-Committee on Stock
Assessment (SCSA). 27 September 2010.
Guillen J., Maynou F., Floros C., Sampson D., Conides A., Kapiris K. (2012). A
bio-economic evaluation of the potential for establishing a commercial fishery
on two newly developed stocks: The Ionian red shrimp fishery. Scientia Marina,
76 (3). doi:10.3989/scimar.03434.07I.
Hammond R.D., Naylor E. (1977). Effects of dusk and dawn on locomotor activity
rhythms in the Norway lobster Nephrops norvegicus. Mar. Biol., 39: 253-260.
Hassan H. (1983). Distribution of penaeid larvae in the coastal waters of
Pakistan. Ph.D. thesis, University of Karachi, Pakistan. pp. 288.
Heldt H. (1938). La reproduction chez les Crustacés décapodes de la famille des
Pénéides. Inst. Océanogr., 18: 1-206.
Heldt H., Heldt J.H. (1954). Les crustacés comestibles des mers tunisiennes et
leur pêche. Annales Sta. Océanogr. Salammbô, 9: 3 – 16.
Heldt J. (1955). Contribution a l’etude e la biologie des crevettes peneides
Aristaeomorpha foliacea (Risso) et Aristeus antennatus (Risso). Bull. Soc. Des
Sc. Tunisie, VIII (1-2): 1-29.
Heldt J.H. (1932). Sur quelques différences sexuelles (Coloration, Taille,
Rostre) chez deux crevettes tunisiennes : Panaeus caramote Risso et Parapenaeus longirostris Lucas. Bull. Sta. Océanogr. Salammbô, (27): 1 - 25.
Heldt J.H. (1938). La reproduction chez les Crustacés Décapodes de la famille
des Pénéides. Ann. Inst. Océanogr. Paris, 28: 1 – 206.
Heldt J.H. (1954). Contribution à l’étude de la biologie des Pénéides. Bull.
Sta. Océanogr. Salammbô, 47: 5 - 27.
Hepper B.T. (1967). Observation on a crawfish (Palinurus vulgaris Latr.) tagging
experiment off Cornwall in 1966. ICES CM. Shelfish and Benthos Committee, 13:
1-4
Hepper B.T. (1970). Observation on the growth of crawfish, Palinurus vulgaris
Latr. Off the coast of cornwall. ICES CM. Shelfish and Benthos Committee, 9:
1-10.
Holthuis L.B. (1980). Shrimps and prawns of the world. FAO. Fish. Synop. 125
(1), 271 p.
Holthuis L.B. (1987). Invertébrés marins : Les crevettes. In : Fiches FAO d’identification
des espèces pour les besoins de la pêche. Zone 37. Révision. W. Fisher M.L.
Bauchot M. Schneider (eds), FAO. Publ. Vol. 1, Rome, pp : 189-292.
Holthuis L.B. (1991). FAO Species Catalogue, Marine Lobsters of the World – FAO
Fisheries Synopsis N° 125, Vol 13. Rome, FAO: 115-122.
Hunter E. (1999). Biology of the european spiny lobster,
Palinurus elephas (Fabricius,
1787) (Decapoda, Palinuridea). Crustaceana, 72(6): 545-567.
Hunter E., Shackley S.E., Bennett D.B. (1996). Recent studies on the crawfish
Palinurus elephas in South Wales and Cornwall. J. Mar. Biol. Ass. U.K., 76:
963-983.
IMBC, UMBSM, and IRPEM. (1994). Nephrops norvegicus stock variability and
assessment in relation to fishing pressure and environmental factors. Final
report to EC, contract XIV-1/MED/91/003.
IRMA–CNR (1999). Valutazione delle risorse demersali nello Stretto di Sicilia
(Mar Mediterraneo) nell’ambito del piano quadriennale 1996–99: rapporto finale –
vol. 1–2. Gruppo Nazionale Valutazione Risorse Demersali (GRUND), Unità
Operativa n° 11, IRMA–CNR, Mazara del Vallo (TP), Italy: 97pp.
Jarboui O. (2009). Available information on closed season in the MedSudMed and
CopeMed II area Tunisian case study. In: Report of the Sub-regional Working
Group on demersal shared stocks in the MedSudMed and CopeMed II area Mazara del
Vallo (Italy), 16-17 September 2010. Presented to the 9th MedSudMed Coordination
Committee. 24-25 March 2011, Capo Granitola-Mazara del Vallo, Italy.FAO-MedSudMed: CC9/Info06. 46pp.
Jaziri H., Ben Meriem S. (2011) Changement spatio-bathymétrique de la taille de
la crevette, Penaeus kerathurus (Forskål, 1775) par sexe et localisation en
Tunisie. 13 eme Journée des Sciences Marine Tunisie et deuxième rencontre
franco-tunisienne d’Ichtyologie 26-29 Novembre 2011. Communication par poster.
Mahdia, Tunisia, unpublished.
Jimenez S., Cano R., Bayle J., Sanchez-Lizaso J.L. (1996). Las praderas de
Posidonia oceanica (L.) Delile como zona de proteccion de juveniles de especies
de interés comercial. Tomo Extraordinario, 125 Aniversario de la Real Sociedad
Española de Historia Natural: 375-378.
Kapiris K. (2012). Feeding Habits of Both Deep-Water Red Shrimps,
Aristaeomorpha
foliacea and Aristeus antennatus (Decapoda, Aristeidae) in the Ionian Sea (E.
Mediterranean). Food Quality, Kostas
Kapiris (Ed.), ISBN: 978-953-51-0560-2,
InTech.
Karani I., Kitsos M.S., Chartosia N., Koukouras A. (2005). Diet Composition of
the Penaeid Shrimp, Melicertus Kerathurus (Forskål, 1775) (Decapoda, Penaeidae)
in the Aegean Sea. Crustaceana, 78 (4): 385-396.
Karlovac O. (1965). Contribution à la connaissance de la biologie de la
langouste commune (Palinurus elephas Fabr.) Note préliminaire). Rapports et
Procès-Verbaux des Réunions du CIESMM. XVIII: 181-184.
Kirkwood G.P., Aukland R., Zara S.J. (2001). Length Frequency Distribution
Analysis (LFDA), version 5.0. MRAG Ltd, London, U.K.
Knittweis L., Dimech M. (2009). Assessment of nursery and spawning areas of
commercially important species in GSA 15. Presentation given at the 11th session
of the GFCM Sub-Committee on Stock Assessment (SCSA) held on the 30/11/2009 –
03/12/2009 in Malaga, Spain.
Knittweis L., Dimech M. (2011). Malta’s Fisheries Management Plan 2011-2015;
Annex VII – Proposed Fishing Protected Area in International Waters;
Recommendations based on the spatial distribution of commercially important
species in GSA 15.
Knittweis L., Arneri E., Ben Meriem S., Dimech M., Fiorentino F., Gancitano V.,
Jarboui O., Mbarek K.B., Ceriola L. (2013). Stock status and potential yield of
deep water rose shrimp (Parapenaeus longirostris, Lucas 1846) in the
south-central Mediterranean Sea. MedSudMed Technical Documents. No. 28. GCP/RER/010/ITA/MSM-TD-28,
Rome, 2013: 15 pp.
Kosmas K., Maria T.L. (2012) Reproductive biology of the
Melicertus kerathurus (Decapoda: Penaeidae) in Thermaikos Gulf (N. Aegean Sea) : 1-15.
Lamboeuf M., Ben Abdallah A., Zgozi S., Natati A., Mer A., Abdubari A. (1995).
Libyan marine resource assessment. Trawl survey results 1993–1994. FI:DP/LIB/88/009
– FI:GCP/LIB/021/IsDB, FAO, Tripoli–Rome. Technical Briefing Notes, 26: 83pp.
Latrouite D., Noel P. (1997). Pêche de la langouste rouge
Palinurus elephas en
France, éléments pour fixer une taille marchande. ICES CM 1997/BB: 13.
Levi D., Andreoli M.G., Giusto R.M. (1995). First assessment of the rose shrimp,
Parapenaeus longirostris (Lucas 1846), in the central Mediterranean. Fish. Res.,
21: 375-393.
Levi D., Vacchi M. (1988). Macroscopic scale for simple and rapid determination
of sexual maturity in Aristaeomorpha foliacea (Risso, 1826) (Decapoda, Penaeidae).
J. Crust. Biol,. 8 (4): 532-538.
Ligas A., Abella A., Colloca F. (2011). SAC GFCM - Sub-Committee on Stock
Assessment (SCSA). 17 October 2011.
Lipcius R.N., Herrnkind W.F. (1987). Control and coordination of reproduction
and molting in the spiny lobster Palinurus argus. Mar. Bio., 96: 207-214.
Lleonart J., Salat J. (1997). VIT: software for fishery analysis. User's manual.
FAO Computerized Information Series (Fisheries). No. 11. Rome, FAO: 105pp.
Loo L.O., Baden S.P., Ulmestrand M. (1993). Suspension feeding in adult Nephrops
norvegicus (L.) and Homarus gammarus (L.) (Decapoda). Neth. J. Sea Res., 31 (3):
291-297.
Maiorano P., Sion L., Carlucci R., Capezzuto F., Giove A., Costantino G., Panza
M., D’Onghia G., Tursi A. (2010). The demersal faunal assemblage of the
north-western Ionian Sea (central Mediterranean): current knowledge and
perspectives. Chemistry and Ecology, 26 (Suppl.): 219–240.
Marano G., Ungaro N., Marzano M.C., Marsan R. (1998). Le Risorse Demersali
dell'Adriatico pugliese: analisi di una serie storica ('85-'95) relativa ai dati
di cattura e demografia degli stock. Biol. Mar. Medit., 5 (2): 52-67.
Marcias S., Sacco F., Cau A., Cannas R. (2010). Microsatellite markers for
population genetic studies of the giant red shrimp Aristaeomorpha foliacea (Crustacea,
Decapoda). Rapp. Comm. Int. Mer Medit., 39.
Marin J. (1985). Etude de la croissance des Crustaces à partir des données de
marquages-recaptures. Application à la langouste rouge de Corse, Palinurus
elephas Fabricius. Journal du Conseil International pur l'Exploration de la Mer
26, 1-17.
Marin J. (1985). La lanoguste rouge: biologie et exploitation. Pêche maritime,
64: 105-113.
Marin J. (1987). Exploitation, biologie et dynamique du stock de langouste rouge
de Corse, Palinurus elephas, Fabricius 1787. Thesis, University College, Galway.
331 pp.
Matarrese A., D’Onghia G., Tursi A., Maiorano P. (1997). Vulnerabilità e
resilienza in Aristaeomorpha foliacea (Risso, 1827) e Aristeus antennatus
(Risso, 1861) (Crostacei, Decapodi) nel Mar Ionio. S. It. E. Atto, 18: 535-538.
Maurin C. (1962). Etude des fonds chalutables de la Méditerranée occidentale
(écologie et pêche): Résultats des compagnes des navires océanographiques «Président-Théodore-Tissier»
1957 à 1960 et « Thalassa » 1960 et 1961.26, ISTPM. 218 pp.
Maurin C. (1965). Repartition des erevettes profondes au large des cotes de
Sardaigne et de Corse. Rapp. Comm. int. Mer Medit., 18 (2): 175-178
Maurin C., Carries C. (1968). Note préliminaire sur l’alimentation des Crevettes
profondes. Rapp. Comm. Int. Mer Médit., 19(2): 155-156.
Maynou F., Cartes J.E. (2011). Effects of trawling on fish and invertebrates
from deep-sea coral facies of Isidella elongata in the western Mediterranean.
Journal of the Marine Biological Association U.K., 92 (7): 1501-1507.
Maynou F., Sardà F. (1997). Nephrops norvegicus population and morphometrical
characteristics in relation to substrate heterogeneity. Fish. Res., 30: 139
–149.
McKoy J.L., Esterman D.B. (1981). Growth of rock lobsters (Jasus edwardsii) in
the Gisborne region. N. Z. J. Mar. Freshw. Res., 15: 121–136.
MedSudMed (2008). Bibliographic synthesis of information on some target species
in the MedSudMed Project area (central Mediterranean). MedSudMed Technical
Documents 15, Rome: 69 pp.
Mercer J.P. (1973). Studies on spiny lobsters (Crustacea: Decapoda: Palinuridae)
of the west coast of Ireland, with particular reference to Palinurus elephas
Fabricius 1787. Ph.D. Thesis, University College Galway, Ireland, 331 pp.
Minervini R. (1996). Guide des homards, crabes, langoustes, crevettes et autres
crustacés décapodes d’Europe. 287 p. Delachaux et Niestlé.
Missaoui H. (2004). The effects of environmental factors on the distribution of
crustaceans along the Tunisian coast. MedSudMed Technical Documents 2.
Missaoui H., Zaouali J. (1995). Apparition de nouveaux Crustacés dans les pêches
crevettières du golfe de Gabès, Tunisie. Marine Life, 5 (2): 27 - 34.
Morratoupolu Kassimati E. (1973). Distribution and fishing of the lobster
Palinurus vulgaris and Homarus vulgaris in Greek seas. Rapp. Comm. Int. Mer.
Médit., 22(4): 69-70.
Mosbah S., Ghorbel M., Jarboui O., Lenfant P., Verdoit-Jarraya M. (2012).
Application des methodes d’analyses des donnees pour la gestion des ressources
naturelles. Statistique Fonctionelle et Operationelle. Recueil des resumes,
Annee 2011-2012: 3-9.
Mytilineou C., Castro M., Gancho P., Fourtouni A. (1998) Growth studies on
Norway lobster, Nephrops norvegicus (L.), in different areas of the
Mediterranean Sea and the adjacent Atlantic. Sci. Mar., 62 (Supl. 1): 43-60.
Mytilineou C., Castro M., Gancho P., Fourtouni A. (1998). Growth studies on
Norway lobster, Nephrops norvegicus (L.) in different areas of the Mediterranean
Sea and the adjacent Atlantic. Sci. Mar., 62 (1): 43-60.
Nouar A. (1985). Contribution a l’étude de la crevette pénéide
Parapenaeus
longirostris (Lucas, 1846), dans la région d’Alger: écologie, biologie,
exploitation. PhD thesis, Univ. Houari Boumedienne, 136 pp.
Needle C.L. (2003) Survey-based assessments with SURBA. Working Document to the
ICES Working Group on Methods of Fish Stock Assessment, Copenhagen, 29 January
to 5 February 2003.
Newman G.G., Pollock D.E. (1974). Growth of the rock lobster Jasus lalandii and
its relationship to benthos. Mar. Biol., 52: 347–356.
Orsi Relini L., Relini G. (1985). The red shrimp fishery in the Ligrian Sea:
mismanagement or not? FAO Fish. Rep., 336: 99-106.
Orsi Relini L., Relini G. (1989). Reproduction of Nephrops norvegicus L. in
isothermal mediterranean waters. In Reproduction, Genetics and Distributions of
Marine Organisms. Ed. J.S. Ryland, P.A. Tyler, Pub. Olsen e Olsen, Denmark:
153-160.
Orsi Relini L., Zamboni A., Fiorentino F., Massi D. (1998). Reproductive
patterns in Norway lobster Nephrops norvegicus (L.), (Crustacea Decapoda
Nephropidae) of different Mediterranean areas. Sci. Mar., 62 (1): 25-41.
Palero F., Abelló P., Macpherson E., Matthee C.M., Pascual M. (2011). Genetic
Diversity Levels in Fishery-Exploited Spiny Lobsters of the Genus Palinurus (Decapoda:
Achelata).
Palero F., Guerao G., Abelló P. (2008). Morphology of the final stagephyllosoma
larva of Scyllarus pygmaeus (Crustacea: Decapoda: Scyllaridae), identified by DNA
analysis. J. Plank. Res., 30: 483–488.
Palero F., Guerao G., Clark P.F., Abelló P. (2011).
Scyllarus arctus (Crustacea:
Decapoda: Scyllaridae) final stage phyllosoma identified by DNA analysis, with
morphological description. Mar. Biol. Ass. U. K., 91(2): 485-492.
Palero, F., Abello, P., Macpherson, E., Gristina, M., Pascual, M. (2008).
Phylogeography of the European spiny lobster (Palinurus elephas): Influence of
current oceanographical features and historical processes. Molec. Phyl. Evol.,
48: 708-717.
Papaconstantinou C., Kapiris K. (2003). The biology of giant red shrimp (Aristaeomorpha
foliacea) at an unexploited fishing ground in the Grek Ionian Sea. Fish. Res.,
62: 37-51.
Pauly D. (1980). On the interrelationships between natural mortality, growth
parameters and mean environmental temperature in 175 fish stocks. J. Cons. CIEM,
39 (3):175-192.
Pauly, D. (1987) A review of the ELEFAN system for analysis of length-frequency
data in fish and aquatic invertebrates. In: D. Pauly and G.R. Morgan (eds.)
Length-Based methods in fisheries researches.
Proc. Int. Conference on the
theory and application of length-based methods for stock assessment, 11-16
February 1985, Mazara del Vallo, Sicily, Italy, ICLARM Conference Proceedings
13: 7-34.
Pellerito R., Arculeo M., Bonhomme F. (2009) Recent expansion of Northeast
Atlantic and Mediterranean populations of Melicertus (Penaeus) kerathurus (Crustacea:Decapoda).
Fish. Sci.,75:1089–1095. DOI 10.1007/s12562-009-0131-z.
Perdichizzi A., Pirrera L., Micale V., Muglia U., Rinelli P. (2012). A
histological study of ovarian development in the giant red shrimp Aristaeomorpha
foliacea (Crustacea: Decapoda: Aristeidae) from the Southern Tyrrhenian Sea
(Western Mediterranean). Scient. World J. doi: 10.1100/2012/289608.
Peres J.M. (1967). The Mediterranean benthos. Oceanography and Marine Biology:
an Annual Review, 5: 449–533.
Pérez Farfante I., Kensley B. (1997). Penaeoid and sergestoid shrimps and prawns
of the world. Keys and diagnoses for the families and genera. Mém. Mus. Nat.
Hist. Nat., 175: 1-233.
Pipitone C., Badalamenti F., Cuttitta C., D’Anna G., Gristina M. (1994).
Preliminary results on the diet of Aristaeomorpha foliacea in the Sicilian
Strait (Central Mediterranean Sea). Proc. Int. Workshop on life cycles and
fisheries of red shrimps. N.T.R.-I.T.P.P. Spec. Publ., 3: 47-48.
Politou C.Y., Kapiris K., Maiorano P., Capezzuto F., Dokos J. (2004). Deep-Sea
Mediterranean biology, the case of A. foliacea (Risso, 1827) (Crustacea,
Decapoda, Aristeidae). Sci. Mar., 68 (3): 117-127.
Pollock D.E. (1979). Predator-prey relationship between the rock lobster Jasus
lalandii and the mussel Aulocomya ater at Robben Island on the Cape west coast
of Africa. Mar. Biol., 52: 347–356.
Pollock D.E. (1982). The fishery and population dynamics of West Coast rock
lobster related to the environment in the Lambert’s Bay and Port Nolloth areas.
Investigations of the Sea Fisheries Institute of South Africa, 124: 57 pp.
Pollock D.E. (1991). Spiny lobster at Tristan da Cunha, South Atlantic:
inter-island variations in growth and population structure. S. Afr. J. Mar.
Sci., 10: 1–12.
Pollock D.E., De Beyers C.J. (1981). Environment, distribution and growth rates
of West Coast rock-lobster Jasus lalandii (H. Milne Edwards). Trans. Royal Soc.
S. Afr., 44: 379–400.
Quetglas A., Gaamour A., Reñones O., Missaoui H., Zarrouk T., Elabed A., Goñi R.
(2004). Spiny lobster (Palinurus elephas, Fabricius 1787) fisheries in the
western Mediterranean: A comparison of Spanish and Tunisian fisheries. Boll.
Soc. Hist. Nat. Baleares, 47: 63-80.
Ragonese S. (1995). Geographical distribution of
Aristaeomorpha foliacea (Crustacea-Aristeidae)
in the Sicilian Channel (Mediterranean Sea). ICES J. Mar. Sci. Symp., 199:
183-188.
Ragonese S., Abella A., Fiorentino F., Spedicato M.T. (2006). Methods for
estimating the instantaneous rate of natural mortality (M) in fisheries science
with particular reference to the Mediterranean. Biol. Mar. Medit., 13(3):
78-151.
Ragonese S., Andreoli M.G., Bono G., Giusto G.B., Rizzo P., Sinacori G. (2004).
Overview of the available biological information on demersal resources of the
Strait of Sicily. GCP/RER/010/ITA/MSM-TD-02. MedSudMed Technical Documents 2:
67-74.
Ragonese S., Bertolino F., Bianchini M.L. (1997). Biometric relationships of the
red shrimp Aristaeomorpha foliacea (Risso 1827), in the Strait of Sicily (Mediterranean
Sea). Sci. Mar., 61(3): 367-377.
Ragonese S., Bianchini M.L. (1995). Size at sexual maturity in red shrimp
females Aristaeomorpha foliacea, from the Sicilian Channel (Mediterranean Sea). Crustaceana, 68 (1): 73-82.
Ragonese S., Bianchini M.L. (2006). Trawl selectivity trials on the deep-water
rose shrimp (Parapenaeus longirostris) in Sicilian waters. Hydrobiologia, 557:
113–119.
Ragonese S., Giusto G.B. (1998). Deep water occurrence of
Penaeus kerathurus (Forskål,
1775) (Decapoda, Penaeidae) in the Strait of Sicily (central Mediterranean Sea). Crustaceana, 71: 353-357.
Ragonese S., Vitale S., Dimech M., De Santi A. (2012). Growth discontinuity in
males of the deep-water giant red shrimp Aristaeomorpha foliacea in the
Mediterranean Sea. Mar. Ecol., 33: 386–392.
Ragonese S., Zagra M., Di Stefano L., Bianchini M.L. (2001). Effect of codend
mesh size on the performance of the deep-water bottom trawl used in the red
shrimp fishery in the Strait of Sicily (Mediterranean Sea). Hydrobiologia, 449:
279-291.
Rainer S.F. (1992). Diet of prawns from the continental slope of North-Western
Australia. Bull. Mar. Sci., 50(2): 258-274.
Rao P.V. (1972). Seasonal abundance of larvae and post-larvae of the
Commercially important penaeid prawns in the inshore Waters of Cochin. Ind. J.
Fish., 19 (1-2): 86-96.
Rawag A.A., Haddoud D.A., Zgozi S.W. (2004). Commercial demersal marine species
of Libya. GCP/RER/010/ITA/MSM-TD-02. MedSudMed Technical Document, 2: 75-81.
Relini G., Bertrand J., Zamboni A. (eds.) (1999). Synthesis of the knowledge on
bottom fishery resources in Central Mediterranean (Italy and Corsica). Biol.
Mar. Medit., 6 (1).
Relini M., Torchia G. (1998). Unexpected displacement of a spiny lobster in the Ligurian sea. Biol. Mar. Medit., 5: 641-643.
Rikhter V.A., Efanov V.N. (1976). On one of the approaches to estimation of
natural mortality of fish populations. ICNAF Res.Doc., 79/VI/8 : 12pp.
Rjeibi O. (2012). Biologie et dynamique des populations de la langouste rouge
Palinurus elephas pêchée sur les côtes tunisiennes. Thèse de doctorat de l’Institut
National Agronomique de Tunisie spécialité Halieutique, Tunisie. 281 pp.
Rjeibi O., Gaamour A., Missaoui H. (2010). Kinetics of ooegenesis and spawning
strategy of the red spiny lobster Palinurus elephas. J. Crust. Biol., 30:
401-412.
Rjeibi O., Gaamour A., Missaoui H. (2011). Etude de la croissance de la
langouste rouge, Palinurus elephas dans les eaux tunisiennes. Bull. Inst. Natn.
Scien. Tech. Mer de Salammbô, vol 38.
Rjeibi O., Gaamour A., Rjeibi O., Missaoui H. (2010). Kinetics of ooegenesis and
spawning strategy of the red spiny lobster Palinurus elephas. J. Crust. Biol.,
30: 401-412.
SAMED (2002). Stock Assessment in the Mediterranean. European Commission,
Directorate-General XIV, Project 99/047, Final Report.
Sardà F., Lleonart J., Cartes J.E. (1998). An analysis of the population
dynamics of Nephrops norvegicus (L.) in the Mediterranean Sea. SCI. MAR., 62 (Supl.
1): 135-143.
Scordella G., Lumare F. (2001). Stato delle popolazioni di
Penaeus kerathurus:
valutazione dello sforzo di pesca e della produttivita nelle aree di Termoli
(CB), Lesina (FG) e Frigole (LE). In: Proceedings of the Workshop Stato della
pesca e dinamica di popolazione del gambero mediterraneo Penaeus kerathurus in
alcune aree della costa adriatica, Lecce, 16 Marzo. pp. 20–27.
Secci E., Cuccu D., Follesa M.C., Cau A. (1999). Fishery and tagging of
Palinurus elephas in Sardinian seas. In: Von-Vaupel Klein, J.C., Schram F.R. (eds.)
Fourth International Crustacean Congress Procedings of the Fourth International
Crustacean Congress, Amsterdam. Brill, Leiden, 665-672. Amsterdam. Brill, Leiden., A.A. Balkema/Rotterdam/Brookfield. The Biodiversity Crisis and Crustacea.
20-7-1998.
Secci E., Cuccu D., Follesa M.C., Sabatini A., Cau A. (1999). Restocking of
Palinurus elephas (Fabr, 1787) in a Western Central Sardinian area. Biol. Mar.
Medit., 6: 614-616.
Serpil Y., Becer Ozvarol Z.A., Ozvarol Y. (2009). Fisheries and Shrimp Economy,
Some Biological Properties of the Shrimp Metapenaeus monoceros (Fabricus, 1798)
in the Gulf of Antalya (Turkey). J. Anim. Vet. Adv., 8: 2530-2536.
Sheehy M.R.J. (1990a). Widespread occurrence of fluorescent morphological lipofuscin in the crustacean brain. J. Crust. Biol., 10: 613-622.
Sheehy M.R.J. (1990b). Potential of morphological lipofuscin age-pigment as an
index of crustacean age. Mar. Biol. Berlin, 107: 439-442.
Sheehy M.R.J., Cameron E., Marsden G., Mcgrath J. (1995). Age structure of
female giant tiger prawns Penaeus monodon as indicated by neuronal lipofuscin
concentration. Mar. Ecol. Prog. Ser., 117: 59-63.
Sobrino I., Silva C., Sbrana M., Kapiris K. ( 2005). A review of the biology and
fisheries of the deep water rose shrimp, Parapenaeus longirostris, in European
Atlantic and Mediterranean waters (Decapoda, Dendrobranchiata, Penaeidae).
Crustaceana, 78: 1153–1184.
Spedicato M.T., Lembo G., Silecchia T., Carbonara P. (1996). Distribuzione e
biologia di Parapenaeus longirostris nel Tirreno centro-meridionale. Biol. Mar.
Medit., 3(1): 579-581.
STECF (2013). In: Cardinale M., Charef A., Osio C. (eds.) Assessment of
Mediterranean Sea stocks part 1 (STECF-13-22) Luxembourg: Publications Office of
the European Union, ISBN 978-92-79-34645-3, doi:10.2788/36268, 400 pp.
STECF 11-14 (2011). Assessment of Mediterranean Sea stocks – part 2. Report of
the Scientific, Technical and Economic Committee for Fisheries. JRC scientific
and policy reports, edited by Cardinale M., Raetz J.-J., Charef A., EUR 25053
EN. 611 pp.
STECF 12-19 (2012). Assessment of Mediterranean Sea stocks – part 1. Report of
the Scientific, Technical and Economic Committee for Fisheries. JRC scientific
and policy reports, edited by Cardinale M., Osio G.C., Charef A., pending EUR
number. 502 pp.
Symonds D.J. (1972). The fishery for the Norway lobster,
Nephrops norvegicus
(L.), off the north-east coast of England. Fishery Investigations, Series II,
27(3): 35pp..
Tabka-Belkhodja S. (2004). Contribution à l’étude écobiologique de la crevette
blanche Metapenaeus monoceros (Fabricius) dans le golfe de Gabès. Mastère. INAT.
68 p.
Taylor C.C. (1959). Temperature and growth. J. Conseil. Explor. Mer., 25 (1):
93-101.
Tidu C., Sardá R., Pinna M., Cannas A., Meloni M.F., Lecca E., Savarino R.
(2004). Morphometric relationships of the European spiny lobster Palinurus
elephas from the northwestern Sardinia. Fish. Res., 69: 371-379.
Tsagarakis K., Palialexis A., andVassilopoulou V. (2013). Mediterranean fishery
discards: reviewof the existing knowledge. – ICES Journal of Marine Science, 71
(5): 1219-1234. doi: 10.1093/icesjms/fst074.
Tuck I.D., Chapman C.J., Atkinson R.J.A. (1997). Population biology of the
Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland-I:
Growth and density. ICES J. Mar. Sci., 54: 125-135.
Turkmen G, Yilmazyerli H. (2006). Some biological aspects of
Melicertus
kerathurus (Forskål, 1775) (Decapoda, Penaeidae) Inhabiting Izmir Bay (Aegean
Sea) Turkey. Crustaceana, 79(5): 583-591.
Turkmen G., Ozden O., Parug S., Akalin S. (2007). Some Reproductive Aspects of
Melicertus kerathurus (Forskål, 1775) (Decapoda, Penaeidae) Inhabiting the
Gulluk Bay (Aegean Sea) of Turkey. J. App. Biol. Sci., 1(1): 69-74, 2007.
Tursi A., Matarrese A., D’Onghia G., Maiorano P., Mastrotataro F., Balanini M.,
Panza M. (1999). Parapenaeus longirostris. In: Relini G., Bertrand J.A., Zamboni
A. (eds.) Synthesis of knowledge on bottom fishery resources in central
Mediterranean (Italy and Corsica). Biol. Mar. Medit., 6(1): 541-553.
Ungaro N., Marano G., Marsan R., Pastorelli A.M. (1999). On the reproduction of
Nephrops norvegicus (L.) in the Southern Adriatic Sea (Mediterranean Sea): sex
ratio, maturity length and potential fecundity. Crustacean Issues, 12: 553-561.
Véron G. (1995). Organisation et classification du règne animal. Nathan : 128
pp.
Vila Y., Medina A., Megina C., Ramos F., Sobrino I. (2000). Quantification of
the agepigment lipofuscin in brains of known-age, pond-reared prawns Penaeus
japonicus (Crustacea, Decapoda). Journ. exp. Zool., 286: 120-130.
Vitale S., Cannizzaro L., Lumare F., Arculeo M., Mazzola S. (2013). Restocking
trial of Melicertus kerathurus (Decapoda, Penaeidae) in the shallow coastal
waters of southwestern Sicily (Mediterranean Sea). Rapp. Comm. Int. Mar Medit.
2013.
Vitale S., Cannizzaro L., Bono G., Beltrano A.M., Milazzo A., Norrito G. (2006).
Catch composition of Decapoda crustaceans from trawl fishery catches in the
Central Mediterranean Sea. J. Coast. Res., Special Issue, 39 (1): 1798-1800.
Vitale S., Cannizzaro L., Lumare L., Mazzola S. (2010). Population parameters of
Melicertus kerathurus (Decapoda, Penaeidae) in Southwest Sicilian shallow waters
(Mediterranean Sea) using Length-Frequency Analysis. Crustaceana, 83 (8):
997-1007. DOI:10.1163/001121610X510615.
Wadie W.F., Abdel Razek F.A. (1985). The effect of damming on the shrimp
population in the south-eastern part of the Mediterranean Sea. Fish. Res., 3:
323–335.
Wilson D.P. (1949). Notes from the Plymouth aquarium, an apparent example of
learning in Palinurus vulgaris Latreille. J. Mar. Biol. Ass. U. K., 28: 347.
Zimmer-Faust R.K., Tyre J.E. Case J.F. (1985). Chemical attraction causing
aggregation in the spiny lobster, Panulirus interruptus (Randall), and its
probable ecological significance. Biol. Bull., 169: 106-l 18.
Zitari-Chatti R., Chatti N., Elouaer A., Said K. (2008). Genetic variation and
population structure of the camarote prawn Penaeus kerathurus (Forskål) from the
eastern and western Mediterranean coasts in Tunisia. Aquac. Res., 39: 70–76.
Zouari M., (1984). Etude descriptive de la pêche à la crevette au filet tramail
fil 40.000 dans la région de Gabès. Rapp. Doc. Inst. natn. scient. tech.
Océanogr. Pêche Salammbô, 2: 31 - 45.
|